习惯被忽略的Poly(A),悄悄成为高分文章的新阵地!
您可能一直没有留意,大多数真核生物的mRNA 3’末端都有由几十个到几百个不等的A碱基组成的poly(A)尾巴。在目前的mRNA测序中,它们的信息一直被忽略。事实上,它们对于mRNA的稳定性和蛋白翻译非常重要,一些常见生理生化途径可能都会受到poly(A)长度的调控作用。
在以下这些常见的场景中,都不难发现poly(A)长度的身影——
1.胚胎发育。在发育初期,胚胎需要启动大量的mRNA来进行快速的分裂和分化。而这些mRNA却是来自妈妈的馈赠——卵子中“封存”了高达50%的基因组的母体mRNA。而这些被“封存”的mRNA,直到早期胚胎时,才打开封印,突然大量启动翻译。
2.免疫反应。急性炎症是对感染和损伤的“快速而有力”的免疫反应。然而,常规的基因转录、转运过程太耗时了。在没有病原入侵的时候,机体也会预先“封存”一批mRNA,一旦需要立即打开封印,实现快速而有力的免疫反应。
3.记忆转化。在短期记忆转化为长期记忆的过程中,大脑内部会发生一些改变。在细胞水平上,表现为神经元结构和功能方面的改变,如形成一些新的突触或增强现有突触联系。在这个过程中,神经元需要在突触处定点合成一些新的蛋白质。表达新蛋白的mRNA由细胞核转录输出,通过长距离运输到达突触。在从细胞核向突触运输的过程中,mRNA一直处于“封存”状态,当它们到达指定位点后才会打开封印,启动定点翻译。
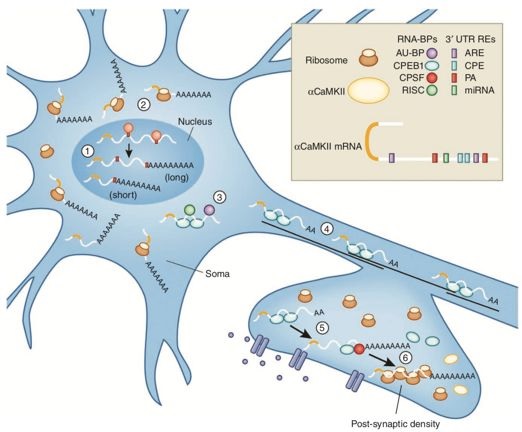
图1 神经元细胞中CaMKII mRNA的循环。CaMKII mRNA在细胞核中经可变剪接产生2种转录本,一种在体细胞中表达,另一种运输到突触中表达。在转录本向突触转运的过程中,mRNA首先在体细胞中被去除poly(A)尾抑制翻译,当mRNA运输到达突触后致密区后,又重新加上poly(A),启动翻译。
在以上过程中,被“封存”的mRNA一直处于表达沉默状态直至打开封印,而打开封印的钥匙就是poly(A)!
什么是poly(A)尾呢?
poly(A)尾即真核生物mRNA的3’尾巴,大多数真核生物的mRNA 3’末端都有由几十个到几百个不等的A碱基组成的poly(A)尾巴。
真核mRNA添加poly(A)尾是一个默认的过程,几乎所有的真核生物mRNA都通过转录后修饰,共获得约250-300个A碱基的poly(A)尾巴。但随后poly(A)的长度在细胞核和细胞质中受到了高度调节,不同长度的poly(A)尾有助于调节成熟mRNA的稳定性、运输和翻译。一旦mRNA到达细胞质,poly(A)尾巴与5′帽协同作用,与翻译起始因子4F(eIF4F)复合物形成稳定的闭环结构,促进翻译起始。poly(A)尾的长度决定了mRNA翻译的程度,poly(A)尾的缩短和延长可以很好地反映poly(A)尾巴在时间和空间上对基因表达的调控。
mRNA脱腺苷酸化,进而脱帽和降解,是细胞核和细胞质中异常或不再需要的转录本沉默机制的第一步,也是必不可少的步骤。但是,mRNA脱腺苷酸化不一定与mRNA降解有关。脱腺苷酸化的mRNA也可以是稳定的,但在翻译上却是沉默的。在许多情况下,这些翻译沉默的转录本会在细胞的特定部位积聚,然后在需要其编码蛋白质的确切时间和特定位置,通过延长poly(A)尾巴将其重新激活。
poly(A)尾巴长度及修饰的变化与许多生理和病理过程有关,胚胎发育、免疫反应和学习与记忆是目前研究最多的跟poly(A)相关的生理过程。
poly(A)测序研究
在目前常规的转录组测序中,poly(A)只是作为反转录引物结合位点,反转录后只保留约20个poly(A)碱基,poly(A)本身的信息一直是被忽略的。
2014年基于短读长测序的poly(A)检测方法的诞生,为基于高通量测序的poly(A)研究提供了可能,目前已有一系列高水平poly(A)研究文章发表。
poly(A)文章一

文章题目:poly(A)尾中散在分布的鸟苷酸(G)可抑制poly(A)结合蛋白(pAB)的结合从而降低mRNA的翻译效率[1]
发表杂志:Genome Biology(IF13.1),2019
发表单位:中国科学院遗传与发育生物学研究所、植物研究所、美国宾夕法尼亚大学
研究方法:TAIL-seq、ribo-seq、mRNA稳定性检测等
poly(A)文章二

文章题目:poly(A)尾的尿酸化促进脊椎动物胚胎发育中母体mRNA的程序性清除[2]
发表杂志:Molecular cell(IF14.25),2018
发表单位:韩国基础科学研究所、首尔国立大学
研究方法:TAIL-seq、ribo-seq等
poly(A)文章三

文章题目:poly(A)尾分析揭示胚胎发育早期到发育成熟过程中翻译调控机制的转变[3]
发表杂志:Nature(IF41.577),2014
发表单位:美国麻省理工学院霍华德·休斯医学研究所、怀特黑德生物医学研究所、麻省理工学院生物系、哈佛-麻省理工医疗科技学院
研究方法:pAL-seq、ribo-seq等
以上几篇文章研究了不同物种poly(A)长度及修饰的变化与翻译的密切关系,以及poly(A)尾的调控在不同生理过程中的重要作用,为基因表达调控研究开辟了新的方向。但受测序技术发展的限制,这几篇文章还都是采用短读长测序的方法。
短读长poly(A)测序受平台限制,有其固有的缺陷:
1. 读长短:短读长poly(A)受读长限制,最长仅能检测230nt左右的poly(A)序列;对于poly(A)较长的物种无法准确获得poly(A)长度分布信息(不同物种间poly(A)尾的长度差异很大,如人poly(A)长度平均是250-300nt,酵母平均是70-80 nt,拟南芥平均是51 nt)。
2. 碱基分辨率低:TAIL-seq的同聚物碱基读取的算法不能准确读取poly(A)尾巴内部A之外的碱基组成。
3. 无法将isoform和poly(A)对应起来:同一基因的不同转录本亚型(isoform)可以具有不同长度的poly(A)尾巴,短读长测序由于读长的限制,无法获得准确isoform信息,无法将转录本亚型与其poly(A)尾巴对应起来。
4. 技术门槛高:TAIL-seq方法操作复杂,市面上尚无科技服务公司提供对应的测序和分析服务,技术门槛高。且要求微克级别的其实RNA,难以应用于微量样品或稀有样品。
poly(A) 全长转录组
为了解决以上问题,华大基因基于pacBio平台自主开发了全长poly(A)研究产品——poly(A)全长转录组。基于pacBio平台的poly(A)测序有短读长平台和ONT平台无法比拟的优势,主要表现在以下方面:
1. 测一得二:poly(A) 全长转录组测序除了全长mRNA序列,还可以得到全长poly(A)序列,加量不加价;可以从RNA整体水平上研究poly(A)长度与isoform及非编码区的关系。
2. 可得到全长的poly(A)信息:基于pacBio平台的poly(A)测序可以直接得到全长poly(A)信息(从几十到几百个碱基不等),满足不同物种不同长度的poly(A)研究需求。
3. 准确度高:poly(A)全长转录组基于Sequel平台CCS测序,可得到准确的poly(A)长度和碱基信息,准确度高;基于短读长平台和Nanopore平台的poly(A)测序都需要通过算法来估算长度,对长度和碱基检测准确度不如poly(A)全长转录组测序。
4. 样本量要求低:poly(A)全长转录组样本要求和常规全长转录组一致,起始样本量可低至1μg总RNA;ONT直接RNA测序对样本量要求很高,需要500ng mRNA(约25μg总RNA)。
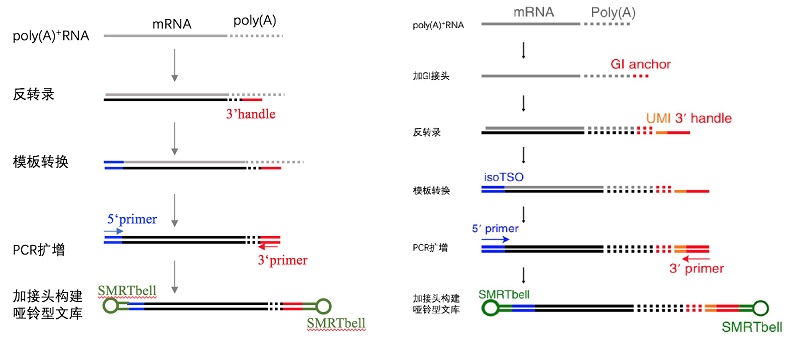
图2 常规全长转录组和poly(A)全长转录组建库方法比较[4]。左图,常规全长转录组,建库过程中以poly(A)为反转录引物结合区域,反转录后只保留约20个poly(A)碱基;右图,poly(A)全长转录组,反转录后保留了全长的poly(A)序列。
作为目前市场上仅有的可以提供poly(A)测序的科技服务公司,华大基因已和客户合作完成了多项poly(A)研究,相关文章也正在发表途中。期待和您一起去探索poly(A)研究的新领域。
参考文献:
1. Zhao T, Huan Q, Sun J, et al. Impact of poly (A)-tail G-content on Arabidopsis pAB binding and their role in enhancing translational efficiency[J]. Genome biology, 2019, 20(1): 1-12.
2. Chang H, Yeo J, Kim J, et al. Terminal uridylyltransferases execute programmed clearance of maternal transcriptome in vertebrate embryos[J]. Molecular cell, 2018, 70(1): 72-82. e7.
3. Subtelny A O, Eichhorn S W, Chen G R, et al. poly (A)-tail profiling reveals an embryonic switch in translational control[J]. Nature, 2014, 508(7494): 66.
4. Legnini, I., Alles, J., Karaiskos, N. et al. FLAM-seq: full-length mRNA sequencing reveals principles of poly(A) tail length control. Nat Methods 16, 879–886 (2019) .































