拟南芥去甲基化酶ALKBH10调控开花通路 | 案例解析

论文标题:ALKBH10B Is an RNA N6-Methyladenosine Demethylase Affecting Arabidopsis Floral Transition
刊登日期:2017年12月
发表杂志:plant Cell
影响因子:8.68
研究机构:北京大学贾桂芳/何川课题组
前言
目前国内外大部分RNA甲基化研究都集中在基础生物学与医学方向,而关于植物RNA甲基化的研究仍然偏少。2014年何川/贾桂芳课题组在Nature Communications上发表研究,利用m6A-seq首次对拟南芥RNA甲基化进行了报道。另外中科院北京基因组研究所以及西北农林科技大学也分别在RNA Biology及Genome Biology上利用m6A-seq对水稻和拟南芥的不同组织部位进行研究。
拟南芥中已鉴定了大量RNA甲基化酶,如METTL3同源基因MTA、METTL14同源基因MTB、WTAp同源基因FIp37以及KIAA1429同源基因Virilizer。去甲基化酶包括ALKBH9A, ALKBH9B, ALKBH9C, ALKBH10A以及ALKBH10B等。其中MTA介导拟南芥胚胎后的转录调控,FIp37与MTA互作,是甲基化酶复合物重要组成部分。
在本文中,贾桂芳课题组利用拟南芥ALKBH10b(At4g02940)突变体研究去甲基化酶如何对拟南芥开花早期的成花转变进行调控。结果显示,ALKBH10b通过介导FT, SpL3和SpL9的去甲基化修饰,并且证实幼年期→成年期关键分子miR-156不参与调控该通路,而是ALKBH10b特异性调控拟南芥成花转变过程。最后论文通过RNA甲基化测序m6A-seq挖掘了1000多个基因发生了差异甲基化修饰,这些基因直接或间接受到了ALKBH10b的影响。
论文框架与流程
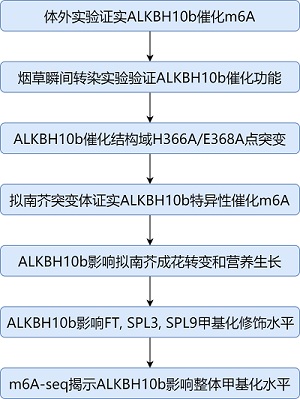
体外实验证实ALKBH10b行使去甲基化作用
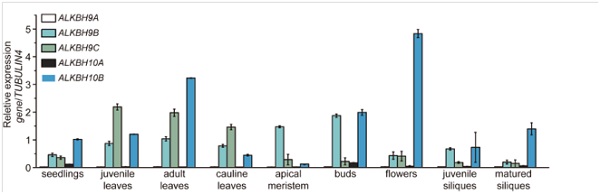
作者先使用BLAST算法,把人源ALKBH5酶与拟南芥的几种酶进行同源序列比对分析,列出了五个候选酶:ALKBH9A, ALKBH9B, ALKBH9C, ALKBH10A和ALKBH10b。结果显示,ALKBH10b在拟南芥几乎所有组织中表达丰度都很高,而且在花期表达量最高。ALKBH9B和ALKBH9C这两种酶的丰度紧接其后。
作者在下一步使用ALKBH9B和ALKBH9C两种拟南芥突变体来验证m6A修饰水平,结果表明mRNA甲基化水平并无明显变化。所以作者选定ALKBH10b作为后续的研究目标。
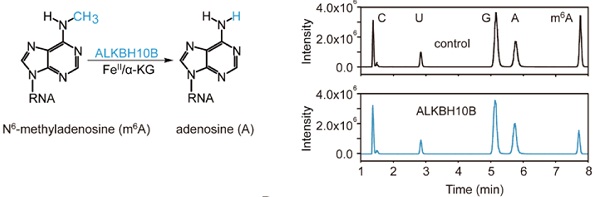
接下来作者分别结合使用了烟草瞬间转染实验以及纯化后的ALKBH10b来研究去甲基化酶在体外的修饰现象。LC-MS/MS分析表明,m6A甲基化水平在体外降低了50%。

最后作者将ALKBH10b行使催化活性关键结构域上的氨基酸残基H366A/E368A进行突变后,ALKBH10b的去甲基化修饰活性被破坏。此外作者还比较了人源ALKBH5在同等条件下与ALKBH10b是否在催化能力上相似。结果表明,ALKBH10b比ALKBH5在催化ssRNA上活性略低,而具有茎环结构的structured RNA两者并无明显差别。另外ALKBH10b只对m6A有特异性去甲基化催化功能,而m1A则没有催化功能。
带有m6A甲基化修饰的mRNA为ALKBH10b的底物
为了在拟南芥体内验证ALKBH10b的功能,作者构建了2个突变体:alkbh10b-1(ALKBH10B FeII结合位点前外显子插入T-DNA)和alkbh10b-2(ALKBH10B预测α-KG结合位点前内含子插入T-DNA)。RT-pCR实验表明alkbh10b-1几乎不产生任何ALLBH10b的转录本,而alkbh10b-2只产生不到5%的转录本。
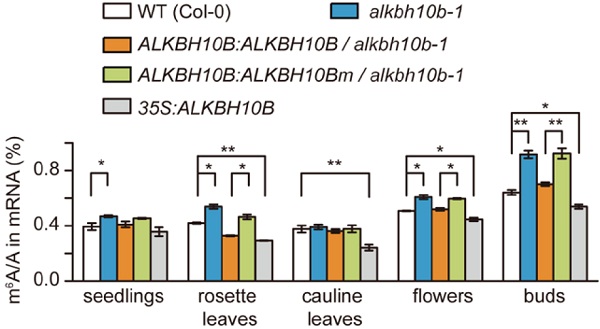
当去甲基化酶ALKBH10b催化功能被破坏后,RNA整体的m6A/A的比例开始升高(即甲基化水平上升)。接下来作者进行补救实验(rescue)对alkbh10b-1和alkbh10b-2进行了ALKBH10b的过表达实验,拟南芥表型得到补救恢复到野生型但alkbh10b-1补救效果比alkbh10b-2要更优。在野生型中过表达ALKBH10b(35S),甲基化水平也是显著下降。
作者对其他几种甲基化转移酶如MTA、MTB、FIp37等进行了qpCR验证,发现并无明显差异。表明ALKBH10b直接参与RNA的去甲基化修饰,使得拟南芥m6A水平发生变化且与甲基化转移酶无关。
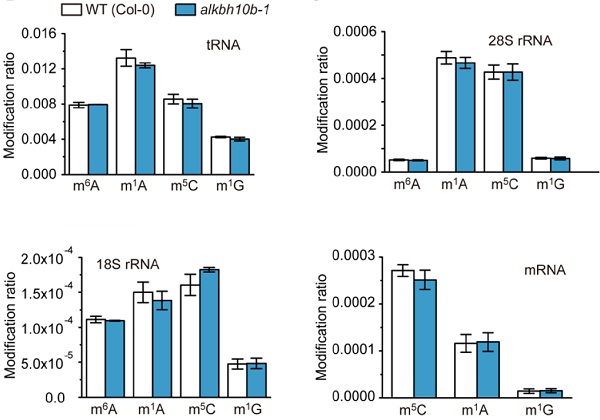
已经有大量的研究,尤其是动物中,核糖体RNA、tRNA等也存在可逆化甲基化修饰和去甲基化修饰。但是最后作者在拟南芥中证实,rRNA和tRNA并不受ALKBH10b的调控,且ALKBH10b也不会对其他类型的碱基修饰如m1A、m5C、m1G等有去甲基化的修饰作用。
所以ALKBH10b的底物不具有广谱性,既不会对rRNA、tRNA上的m6A有去甲基化修饰作用,也不会对其他修饰如m1A等有催化功能。
ALKBH10b影响拟南芥成花转变和营养生长
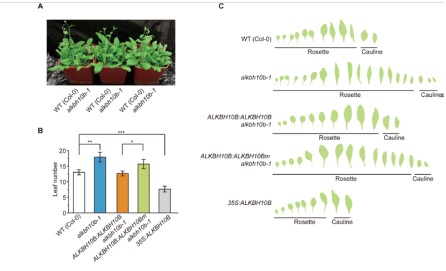
作者对alkbh10b-1突变体进行长时间观察后发现,野生型在展开第12片真叶(true leaves)后开花,而alkbh10b-1则花蕾绽放前额外多出了7片真叶。相反的是,在野生型中过表达ALKBH10b在产生7片真叶后提早进入花期。在alkbh10b-1突变体中进行ALKBH10b补救实验,可对alkbh10b-1中原本ALLBH10b产生缺陷的表型进行补救,从而恢复到野生型的表型水平。然后,针对alkbh10b-1进行ALKBH10b H366A/E368A(即催化结构域被突变的ALKBH10bm)的补救后,发现表型依旧没有恢复到野生型水平。最后作者得出结论ALKBH10b功能缺失后成花转变延后,营养生长受到抑制。
ALKBH10b介导去甲基化修饰提升开花关键基因转录水平
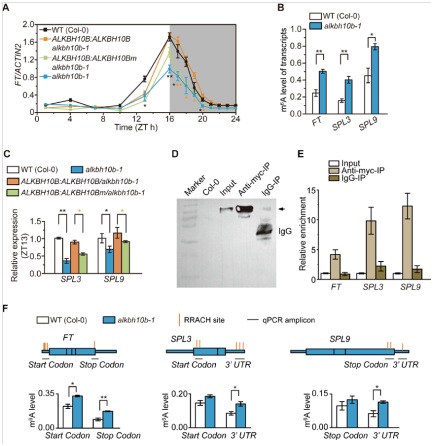
已知FT基因(Flowering Locus T)参与拟南芥叶片光周期、叶片春化、赤霉素和衰老通路。在alkbh10b-1突变体中,FT基因表达量显著降低,花期延后。说明ALKBH10b能够调节FT基因的丰度最终影响成花转变。为了进一步证实m6A甲基化修饰参与FT表达,作用使用了m6A-RIp-qpCR技术对FT甲基化水平进行检测后发现,超过50%的FT在alkbh10b-1中发生了m6A甲基化修饰,而野生型中m6A修饰水平不到30%。此外,与FT基因相关的SpL3和SpL9在alkbh10b-1突变体中甲基化水平也是显著提升,转录水平显著下降。通过m6A-RIp-qpCR技术对片段化的mRNA进行检测后发现,FT基因peak主要位于起始及终止密码子,SpL3及SpL9的peak主要位于3’ UTR区域。所以结合m6A-RIp-qpCR实验不仅证明了ALKBH10b直接参与介导FT、SpL3和SpL9的去甲基化修饰,同时表明去甲基化修饰是一种转录后调控基因表达水平的一种方式。
ALKBH10b维持FT, SpL3和SpL9的稳定性并影响成花转变
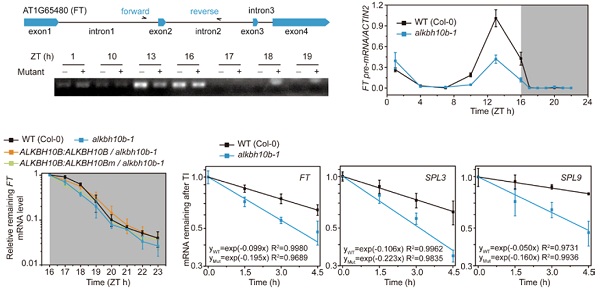
在哺乳动物中,先前已有大量实验证实YTHDF2作为reader可以对发生甲基化的mRNA进行降解。为了在拟南芥中证实这种猜想,作者开展了两项独立实验来对FT基因在整个生命周期中的降解情况进行实时监控:a. 在转录水平最高的白天Z16期前使用转录抑制剂放线菌素D(actinomycin D)监控FT的转录水平;b. 在夜晚Z16后检测FT的降解水平。两种方法同时证实在alkbh10b-1突变体中FT降解速度比野生型拟南芥更快。这个现象表明,当拟南芥中甲基化水平提升时,mRNA会加速降解。当在alkbh10b-1中进行ALKBH10b过表达时,FT甲基化水平降低,mRNA维持稳定性(即不被快速降解)。
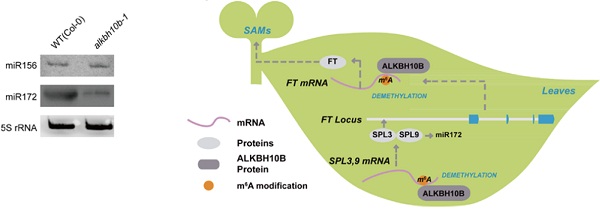
已知miR-156调控SpL基因。宾夕法尼亚大学的Scott教授与浙江农林大学的吴刚教授首次发现miR-156是控制植物幼年向成年阶段转变的主控因子,并揭示了控制该发育进程的信号通道。但是在本次实验中,作者证实miR-156并不参与调控SpL3和SpL9。这表明SpL3和SpL9转录水平下降与miR-156没有关联。所有结果均表明去甲基化酶ALKBH10b能够维持FT、SpL3和SpL9 mRNA的稳定性。
FT、SpL3和SpL9的高甲基化水平能够加速降解,而FT的缺失导致花期延后。在叶片中,SpL9能够激活miR-172从而调控花期,而SpL3能够直接与FT的启动子相结合。在alkbh10b-1中miR-172成熟体和前体数量均显著下降。所以SpL3和SpL9因为高甲基化修饰导致转录水平下降,使得FT和miR-172受到抑制,从而导致成花转变延后。
ALKBH10b缺失导致拟南芥整体m6A修饰水平降低
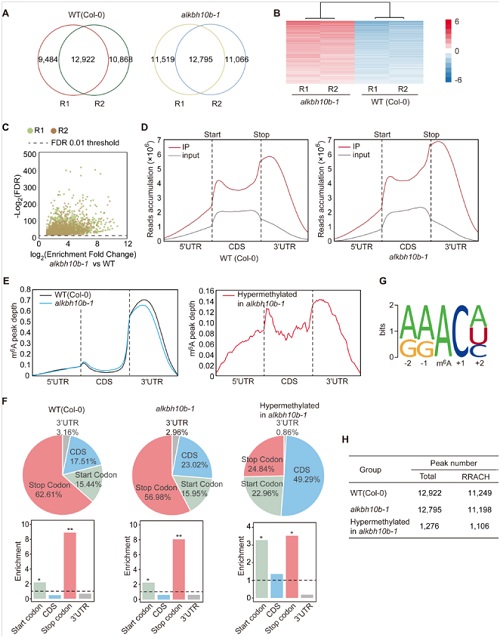
作者使用了m6A-seq和转录组测序等手段,通过多组学联合分析最终发现在野生型拟南芥中的9210多个基因挖掘到了12922多个peak;alkbh10b-1突变体的8000多个基因上挖掘12975个peak。差异分析表明,alkbh10b-1中共鉴定到1190个基因存在1276过甲基化修饰peak。m6A修饰主要位于TSS、TES以及3’ UTR和5’ UTR附近。本次测序结果,m6A peak基本集中在RRACH motif内。GO功能富集分析表明,差异基因基本与器官组织发育、生殖成熟及开花转换等功能相关。这个结果与之前alkbh10b-1表型一致。
总结
这篇文章详细阐述了去甲基化酶介导的m6A修饰可以维持与花期相关基因的mRNA稳定性,从而影响下游通路基因功能,为植物发育的研究提供了新思路。































