Nature子刊,Cell子刊两篇文章报道紫外光调控的光形态建成新突破
光形态建成是植物幼苗出土后由黑暗环境转入光照条件下的必经发育过程。正常的UV-B光形态建成不仅促进植物生物量的增加,也能够增强植物抵御UV-B损伤与病虫害侵袭的能力,但具体信号传递机制并不清楚。
中科院上海生命科学院植物生理生态研究所的研究人员发现UVR8能通过直接结合BES1和BIM1调控光形态建成,这研究成果公布在Developmental Cell杂志上。
同时这一研究组也发现UVR8能结合WRKY36,调控HY5表达和下胚轴伸长,这一研究成果公布在Nature plants杂志上。
这些发现是紫外光调控的光形态建成研究的突破性进展,拓展了人们对紫外光调控植物发育的了解,对推动 UV-B 光能源在农业生产中科学有效的应用具有重要意义。
紫外光UV-B可以作为信号调控植物发育,如调控光形态建成(抑制植物伸长、促进子叶张开以利于光合作用),促进类黄酮和花青素积累(防晒抗逆)等。光形态建成是植物幼苗出土后由黑暗环境转入光照条件下的必经发育过程。正常的UV-B光形态建成不仅促进植物生物量的增加,也能够增强植物抵御UV-B损伤与病虫害侵袭的能力。UVR8(UV RESISTANCE LOUCS 8)是植物中紫外光UV-B受体,是所有光受体中最后被发现的,直到2011年才被报道,具体信号传递机制并不清楚,因此其信号传递机制研究不仅具有重要理论意义,还将为作物育种提供理论依据。
上海生命科学院植物生理生态研究所刘宏涛研究组的研究发现,UVR8能够直接结合内源激素油菜素甾醇(BR)信号转导中的关键转录因子BIM1、BES1,以及功能未知的转录因子WRKY36。UVR8蛋白直接结合BES1及WRKY36转录因子并抑制它们的DNA结合活性进而抑制下游基因表达及伸长,实现光形态建成调控。
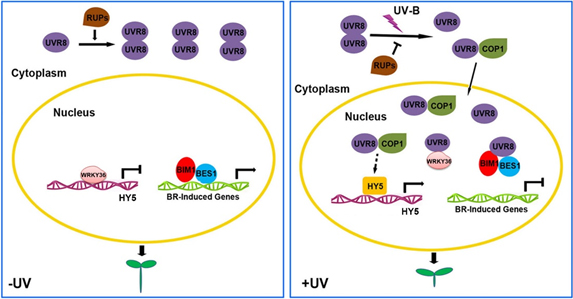 紫外光UV-B调控光形态建成模型图
紫外光UV-B调控光形态建成模型图
UV-B一方面通过抑制植物内源激素油菜素甾醇促进的生长而实现对伸长的抑制,另一方面则通过解除WRKY36对HY5表达的抑制而抑制伸长。UV-B促进的UVR8-BES1/BIM1及UVR8-WRKY36结合是光受体UVR8信号转导的早期机制,也是两条全新的UV-B信号通路。同时UVR8-BES1也是外源光信号和内源油菜素甾醇信号协同调控植物发育的整合点。
这些研究拓展了人们对紫外光调控植物发育的了解,对推动 UV-B 光能源在农业生产中科学有效的应用具有重要意义。
原文标题:
UVR8 Interacts with WRKY36 to Regulate HY5 Transcription and Hypocotyl Elongation in Arabidopsis
UVR8 Interacts with BES1 and BIM1 to Regulate Transcription and photomorphogenesis in Arabidopsis































