浙大学者发文揭示裂殖酵母中染色体倍性增加与中心体数目控制的关系
2021年11月8日,我院何向伟课题组在Journal of Cell Biology在线发表了题为“pcp1/pericentrin controls the SpB number in fission yeast meiosis and ploidy homeostasis”的研究论文。
大部分真核生物主要以单倍体或者二倍体的形式生活,生物体个体的多倍体化在自然界中并不常见。研究表明,多倍体在生物进化过程中发挥着重要作用,现存的物种中有很多在进化过程中都曾存在过多倍体的染色体组型。大量的研究和当前的进化结果告诉我们“Moreis not always better”。比起染色体增加所带来的优势,伴随多倍体所存在的遗传高不稳定性制约着多倍体的形成与发展。但这其中的机制并不清楚。
中心体(在酵母中称为spindlepolebody,SpB)的数量在细胞分裂过程中需要受到严格的控制,以确保在细胞分裂过程中正确组装双极的纺锤体来介导染色体的准确分离。特别的是,有性生殖过程中,为了保证受精卵(接合子)只持有一个中心体然后复制以进行接下来的细胞分裂,精子和卵细胞的中心体需要融合成一个中心体来发挥作用。为此,在配子形成过程中,中心体需要经历一个“减负”过程。例如精子形成过程中的“centrosomereduction”,这个过程主要包括中心体外周物质(pericentriolar material,pCM)的消失。
在酵母的有性杂交时,两个配子的SpB也需要进行融合以保证接合子只持有一个SpB。本研究发现,裂殖酵母杂交时配子SpB的融合以及接下来细胞分裂过程中SpB数量的稳定同样需要pcp1(pCM组分pericentrin的同源蛋白)从SpB上解离。由二倍体杂交而来的多倍体接合子中由于pcp1不能正常的从SpB上解离,会导致SpB融合的阻断,从而导致多倍体接合子在接下来的减数分裂过程中产生多余的SpB以及多极的细胞核分裂,最终导致多余孢子的形成。此外,由二倍体杂交而来的三倍体和四倍体接合子在营养恢复的时候由于异常的SpB数量也不能回到稳定的有丝分裂增殖,不能以多倍体的形式进行有丝分裂传代。同时,通过构建融合蛋白pcp1-ppc89-GFp(ppc89为在SpB上持续定位的SpB组分)在二倍体接合子中模拟pcp1在SpB上的持续定位也充分可以阻断SpB的融合并导致多余SpB的产生,进一步验证了pcp1在有性杂交早期的解离对接合子SpB数量控制的作用。
在多个物种中的研究表明,细胞的四倍体化可以导致肿瘤的形成,这与四倍体细胞的高的遗传不稳定性是密切相关的。普遍认为,肿瘤细胞中多余的中心体都是伴随着细胞的多倍体化而产生的,而且其高的遗传不稳定性与这些多余中心体的存在也密切相关。但其实多倍体化与中心体数目的关系远比想象的复杂,多倍体化和多余中心体单独都可以引发肿瘤形成。本研究在酵母中第一次探讨了染色体倍性增加与中心体数目的关系。进一步表明多倍体化与微管组织中心增加之间的关系在从酵母到哺乳动物的多种生物中是保守的。此外,我们认为微管组织中心(中心体或者SpB)数量的严格控制是多种生物中限制倍性增加的关键因素,从而决定了大多数真核生物的二倍体或单倍体染色体组型。而多倍体高等植物的普遍存在似乎支持了这一猜测。高等植物细胞中不存在中心体,进行无中心体的纺锤体微管组装,对于多倍体化却有很强的容忍度。更重要的是,种间杂交产生的多倍体化是其物种进化的重要方式。
该工作获得科技部973项目和国家自然科学基金委面上项目的资助和支持。博士研究生祝倩是本文的第一作者。
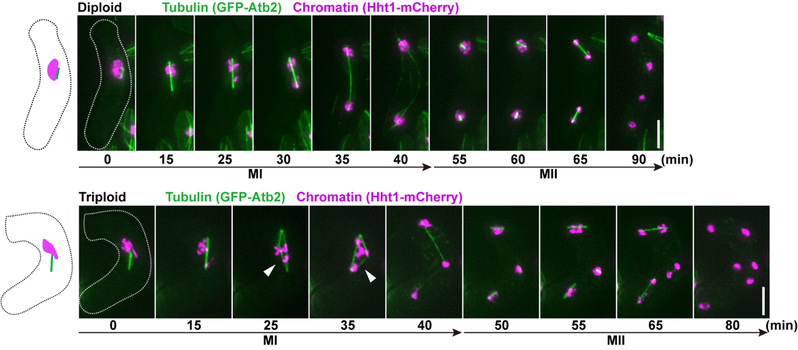
图注:二倍体和多倍体接合子减数分裂过程比较。
如图所示,三倍体接合子减数第一次分裂过程中组装了多极的(三角形)纺锤体,导致多极的染色体分离。
原文链接:https://doi.org/10.1083/jcb.202104099































