施一公连发两篇Cell文章:一步步完成剪接体的拼图
——同日Science杂志也公布了科罗拉多大学丹佛分校Rui Zhao研究组的相似成果
清华大学施一公教授研究组一直致力于捕捉RNA剪接过程中处于不同动态变化的剪接体结构,从而从分子层面阐释RNA剪接的工作机理。
在11月16日公布的Cell杂志上,这一研究组再次发表研究论文:Structure of the post-catalytic Spliceosome from Saccharomyces cerevisiae,公布了一个来自酿酒酵母的p复合物(分辨率为3.6Å)的冷冻电镜结构。两个月前,施一公教授研究组也在Cell杂志上发文,报道了RNA剪接循环中剪接体最后一个状态的高分辨率三维结构,这也是这一研究组今年在Cell杂志上发表的第三篇文章,他们在一步步完成剪接体的拼图。

2015年,施一公研究组率先突破,在世界上首次报道了裂殖酵母剪接体3.6埃的高分辨率结构,首次展示了剪接体催化中心近原子分辨率的结构。自2015年第一个剪接体结构发表以后,施一公研究组相继解析了5个不同状态剪接体复合物的高分辨率结构,分别是酿酒酵母3.8Å的预组装复合物U4/U6.U5 Tri-snRNp、3.5Å的激活状态复合物Bact complex、3.4Å的第一步催化反应后复合物C complex、4.0埃的第二步催化激活状态下的C* complex,以及前文3.5Å的内含子套索剪接体ILS complex的结构。
这个5个不同状态的剪接体基本覆盖了整个剪接通路中从预组装到激活、从发生两步转酯反应到剪接体的解聚的关键催化步骤,呈现了迄今为止最为清晰的剪接体不同工作状态下的结构信息,将RNA剪接领域的发展推向了新的高度。施一公教授因此于不久前也获得未来科学大奖生命医学奖。
相比于原核生物,真核生物的基因表达更为复杂也更为精细。由于真核细胞内的基因是不连续的,需要在细胞核内被转录成前体信使RNA (pre-mRNA)后,通过RNA剪接,不具有翻译功能的内含子(intron)被去除,密码子所在的外显子(exon)被连接,从而得到成熟的、可被翻译成蛋白质的mRNA。RNA剪接是真核生物基因表达调控的重要环节之一,是“中心法则”的关键步骤之一。而负责执行这一过程的是细胞核内一个巨大的且高度动态变化的分子机器——剪接体(spliceosome)。剪接体在真核生物进化中极为保守,对于真核生物维持正常的生命活动至关重要。一个基因转录出的pre-mRNA可以通过RNA剪接形成若干种mRNA,于是极大地丰富了真核生物蛋白质组的多样性。在剪接反应过程中,多种蛋白质-核酸复合物及剪接因子按照高度精确的顺序发生结合和解聚,依次形成预组装复合物U4/U6.U5 tri-snRNp以及至少7个状态的组装剪接体B、Bact、B*、C、C*、p以及ILS复合物。
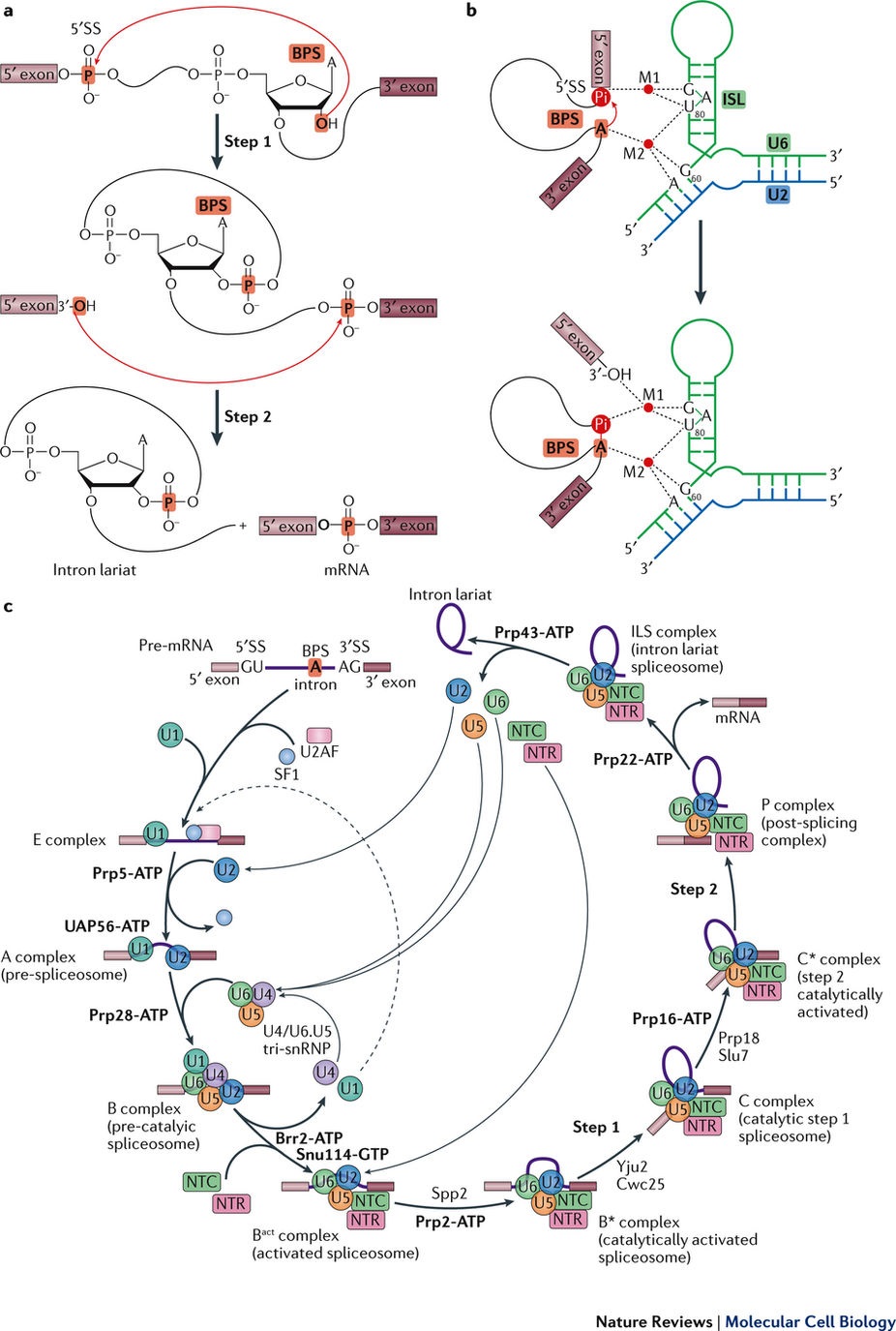
通过之前的努力,已经解析了酿酒酵母或人源的预组装复合物U4/U6.U5 tri-snRNp、催化前复合物B complex、激活状态复合物Bact complex、第一步催化反应复合物C complex、第二步催化激活状态下的C* complex和内含子套索剪接体ILS complex。除了B complex,其它四个状态下的主要的结构特色都与裂殖酵母ILS complex的一致。这样,就只剩下催化激活的复合物B* complex和催化后复合物p complex在结构上未被表征。
在之前研究的基础上,研究人员对36个剪接体蛋白,3个snRNA(U2、U5和U6),1个连接的外显子和1个内含子套索进行了原子建模。最终的结构模型包括了9328个氨基酸酸残基和381个RNA核苷酸,联合起来的分子质量达到了~1.2 MD。如之前预测的一样,p complex的总的组织形式和详细的结构特色与C* complex的非常类似。
在p complex中,步骤II剪接因子prp17, prp18和Slu7仍结合在活性位点附近。剪接因子Cwc21和Cwc22一起稳定了外显子。ATp酶/解螺旋酶prp22位于高度不对称的p complex的一个角落,主要与prp8的Lingker结构域相互作用。连接的外显子锚定在U5 snRNA的loop上,保守的3’-拼接位点(3 ‘SS)的AG二核苷酸则坐落于活性位点。
尽管p complex的结构高度类似于C* complex,该结构仍然在剪接体的活性位点处揭示了意想不到的发现,该发现对我们机制的理解pre-mRNA的拼接具有十分重要的启示作用。同时结构的阐释使得我们理解了剪切体C*到p 和 the p到ILS 的转变机制,因而是完成pre-mRNA剪接循环的必须一步。
相关文章:
清华大学施一公教授最新综述:终于水落石出的原子结构
施一公:当年放弃国外教授职位很多人认为我疯了
清华大学施一公年底再发Science文章:六篇文章把一个故事说完整
原文标题:
Structure of the post-catalytic Spliceosome from Saccharomyces cerevisiae































